KONTROL
GENETIK TERHADAP RESPON IMUN
(Komponen
Sistem Imun, Antibody Diversity (Penyusunan Kembali Genom selama Diferensiasi
Limfosit B; Jalur Alternatif pada
Penyambungan Hasil Transkripsi ))
Ketika substansi asing yang disebut sebagai antigen (misalnya protein selubung virus) memasuki aliran darah
mamalia, maka tubuh mamalia akan memicu mekanisme pertahanan, yaitu respon
imun, yang mengakibatkan sintesis kelompok protein yang penting yaitu antibodi.
Antibodi-antibodi tersebut mengikat antigen dengan spesifikasi dan
memfasilitasi pengeluarannya dari sistem sirkulasi. Para ilmuan telah menemukan
sekuen-sekuan DNA yang mengkode susunan antibodi yang dihasilkan oleh sistem
imun mamalia yang terkait diferensiasi sel-sel penghasil antibodi dengan
terjadinya suatu set baru dari penyusunan kembali genom (rearrangements genome).
1.
Komponen
Sistem Imun
Terdapat tiga tipe sel darah putih yang
berperan dalam respon imun pada vertebrata. Yaitu:
- Limfosit B (disebut sel B karena diproduksi di dalam
sumsum tulang (bone marrow))
- Limfosit T (disebut sel T karena di produksi dalam
kelenjar timus)
- Makrofag
Antibodi-antibodi disintesis oleh limfosit B dan antibodi
ini bisa disekresikan atau tetap terikat pada membran pada permukaan sel B,
bergantung pada kondisinya. Selama respon imun humeral, antibodi-antibodi
mengikat antigen bebas dalam sistem sirkulasi dan mengaglutinasi
antigen-antigen tersebut. Kompleks antibodi-antigen yang dihasilkan kemudian
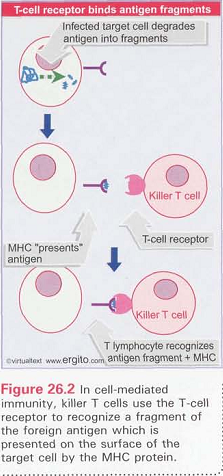
diingesti dan didegradasi oleh makrofag. Limfosit T menengahi respon imun
seluler. Limfosit T mensintesis reseptor-reseptor antigen yang mengenali
antigen pada permukaan sel dan mengenali antigen melalui aktivasi sel-sel T.
Limfosit T yang berbeda menunjukkan cara kerja yang berbeda, akan tetapi secara
umum serangan sel T terhadap sel yang membawa antigen membutuhkan reseptor sel
T yang spesifik dari satu atau lebih reseptor antigen histokompatibilitas (Gardner, 1991).


1.
Berbagai
Kajian Antibodi
Aspek yang paling patut diperhatikan dari respon imun
dari sudut genetik adalah nampaknya varietas antibodi yang bisa disintesis
dalam merespon antigen yang sebelumnya belum diketahui pada hewan. Berapa macam
antigen yang bisa dihasilkan oleh tikus dan manusia belum dapat diketahui bahwa
jumlahya sangat besar, sampai jutaan. Genome manusia lengkap (misalnya satu
dari 23 pasang kromosom manusia) mengandung kurang lebih 3 x 109
pasang nukleotida. Jika semua berada dalam bentuk gen-gen dengan urutan
pengkodean terganggu yang masing-masing panjangnya 1000 pasang nukleotida,
gen tersebut maksimum mengandung 3 juta gen.
2.
Beberapa
Hipotesis Dasar Genetika Keanekaragaman Antibodi
Dasar geenetika mengenai keanekaragaman antibodi
secara umum dapat dikelompokkan menjadi
tiga hipotesis, yaitu:
1. Hipotesis
germ line yang menyatakan bahwa
terdapat germ line yang terpisah
untuk setiap antibodi.
2. Hipotesis
mutasi tubuh, yang menyatakan bahwa terdapat satu atau beberapa germ line spesifik untuk setiap kelas
antibodi, dan keanekaragamannya
disebabkan karena tingginya frekuensi mutasi somatik, yaitu mutasi yang
terjadi pada sel-sel somatik
penghasil antibodi
atau dalam garis sel yang mengarah pada penghasil antibodi.
3. Hipotesis
minigene. Keanekaragaman disebabkan
oleh “suffling” (pengocokan) segmen-segmen kecil beberapa
gen menjadi sejumlah besar kemungkinan kombinasi. Suffing akan terjadi melalui proses rekombinasi pada sel somatik (secara
total ini memerlukan mekanisme untuk menyusun kembali segmen DNA).
Sekarang diketahui bahwa hipotesis minigen menjelaskan
keanekaragaman yang dapat diketahui. Selain itu diketahui pula bahwa mutasi
somatik memberikan kontribusi dalam keanekaragaman. Akhirnya dapat diketahui
bahwa satu segmen dari setiap rantai antibodi ditentukan oleh gen atau segmen
gen yang terdaat pada genome. Dengan demikian kesimpulannya adaah ketiga
hipotesis tersebut adalah benar dalam hal tertentu.
4.
Struktur
Antibodi
Antibodi termasuk kelas protein yang disebut
immunoglobulin. Setiap antibodi
adalah tetramer yang tersusun atas 4 polipeptida, 2 rantai ringan yang identik dan 2
rantai berat yang identik, tergabung oleh ikatan disulfida. Setiap rantai,
berat maupun ringan mempunyai ujung amino daerah variabel,
dimana sekuen asam amino bervariasi di
antara antibodi
spesifik untuk antigen-antigen yang bebeda, dan suatu ujung karboksil daerah konstan,
dimana sekuen asam aminonya sama untuk semua antibodi dari kelas immunoglobulin
tertentu.

Daerah protein yang membawa fungsi khusus disebut domain. Setiap antibodi memiliki 2 domain,
dimana setiap domain dibentuk oleh variable region dari satu rantai ringan dan
satu rantai berat. daerah konstan dari 2 rantai
berat berinteraksi membentuk domain
ketiga yang disebut effector function
domain, yang dapat merespon interaksi
yang sesuai dari antibodi dengan komponen-komponen lain dari sistem imun.
Terdapat 5 kelas antibody yaitu IgM, IgD, IgG, IgE,
IgA. Pengelompokan antibodi tersebut dan fungsinya ditentukan oleh struktur
rantai berat daerah konstan, yaitu struktur dan effector function domainnya. Sebagai contoh antibodi IgD biasanya tetap terikat pada
permukaan sel tempat mereka disintesis, sedangkan antibodi IgG biasanya
disekresikan dan disirkulasikan ke seluruh tubuh melalui aliran darah. Rantai
ringan antibodi mempunyai dua tipe, yaitu kappa dan lambda. Tipe tersebut
ditentukan oleh struktur rantai ringan daerah konstan. Antibodi memiliki
spesifikasi antigen-binding yang sama, yang ditentukan oleh daerah variabel
pada keempat rantai, tetapi fungsi imunoglobinnya berbeda yang ditentukan oleh
daerah konstan pada dua rantai berat. Ketika mempelajari struktur antibodi
dapat dilihat bahwa keanekaragamannya hampir
seluruhnya terletak pada bervariasi
daerah molekul daerah variabel suatu molekul.
Pertanyaan
- Jelaskan hipotesis-hipotesis yang digunakan sebagai
dasar keanekaragaman antibodi!
Jawaban
Dasar geenetika mengenai keanekaragaman antibodi
secara umum dapat dikelompokkan menjadi
tiga hipotesis, yaitu:
- Hipotesis
germ line yang menyatakan bahwa
terdapat germ line yang terpisah
untuk setiap antibodi.
- Hipotesis
mutasi tubuh, yang menyatakan bahwa terdapat satu atau beberapa germ line spesifik untuk setiap
kelas antibodi,
dan keanekaragamannya disebabkan karena tingginya frekuensi mutasi somatik, yaitu mutasi
yang terjadi pada sel-sel somatik penghasil antibodi atau dalam garis
sel yang mengarah pada penghasil antibodi.
- Hipotesis
minigene. Keanekaragaman
disebabkan oleh “suffling” (pengocokan)
segmen-segmen kecil beberapa gen menjadi sejumlah besar kemungkinan
kombinasi. Suffing akan
terjadi melalui proses rekombinasi pada sel somatik (secara total ini
memerlukan mekanisme untuk menyusun kembali segmen DNA).
DAFTAR PUSTAKA
Gardner, E, J.,
Michael J. Simmons, D. Peter Snustad.
1991. Principles of Genetic Eighth Edition.
Lewin, B. 2004. Genes VIII Lewin. United States of America:
Pearson Prentice Hall, PearsonEducation,Inc.















